Störd glutamatreglering kan förklara bristande informationsfiltrering | sid 1137 | 07-04-03
»Trött i hjärnan« osynligt handikapp som kan ge stora problem
Lars Rönnbäck, professor, överläkare i neurologi, Sahlgrenska Universitetssjukhuset/ Sahlgrenska, Göteborg
lars.ronnback(at)neuro.gu.se
Mikael Persson, med lic, doktorand; båda institutionen för neurovetenskap och fysiologi, sektionen för klinisk neurovetenskap och rehabilitering, Sahlgrenska akademin vid Göteborgs universitet
mikael.persson(at)neuro.gu.se
Torsten Olsson, professor, institutionen för signaler och system, Chalmers tekniska högskola, Göteborg
torsten(at)s2.chalmers.se
Sammanfattat
Koncentrations- och minnessvårigheter och mental uttröttbarhet försvårar rehabilitering och återgång till arbete efter stroke och traumatisk hjärnskada. Det är också vanliga symtom vid multipel skleros och andra inflammatoriska sjukdomar.
Mental uttröttbarhet innebär att man kan koncentrera sig en kort stund men sedan måste vila. Liknande uttröttbarhet förekommer också vid långvariga värk- och stresstillstånd, dvs vid tillstånd utan uppenbar nervcellsskada.
Glutamatsignaleringen är av stor betydelse för hjärnans intag och bearbetning av information.
Astrocyterna har viktig roll i reglering av glutamatsignaleringen. Precisionen i glutamatreglering minskar vid skada.
Stort informationsinflöde i kombination med bristande precision i bearbetningen leder på det cellulära planet till förändrade jonkoncentrationer, cellsvullnad och minskad glutamattransmission, vilket kan ge en neurobiologisk förklaring till uttröttbarheten.
Alla blir vi trötta när vi koncentrerat oss länge och intensivt på någon mental uppgift. Det kan handla om faktaspäckade föreläsningar eller tentamensläsning om man är student, att gå igenom och sammanfatta utredningar på jobbet eller att som lärare undervisa i nya moment. Man upplever att man blir »trött i huvudet« eller »trött i hjärnan«. Man behöver göra ett avbrott, kanske vila en stund eller ta en kopp kaffe, för att därefter kunna fortsätta med sina uppgifter. Detta är normalt.
Om vi sovit eller ätit för lite eller om vi är ordentligt förkylda, blir vi lättare uttröttbara med bristande ork. Av erfarenhet vet vi dock att vi, åtminstone delvis, kan »sätta oss över« denna uttröttbarhet. Dessutom vet vi att när vi tagit igen den förlorade sömnen, fått i oss mat eller infektionen gått över är den ökade uttröttbarheten borta.
Sjuklig uttröttbarhet – energin »tar slut«
Mental uttröttbarhet i patologisk eller sjuklig mening yttrar sig i form av mer extrema koncentrationssvårigheter eller nedsatt förmåga att behålla uppmärksamheten över tiden. Man kan säga att energin efter hjärnaktiviteten »tar slut«. Personen kan kanske bara läsa några få sidor i en bok i sträck, och det räcker inte med en kort paus för att vara alert igen. Det behövs regelbundna pauser och ibland ordentlig vila, vilket leder till svårigheter att genomföra även mindre krävande intellektuella uppgifter på rimlig tid.
Om behovet av täta avbrott inte respekteras blir personen utmattad och kan få huvudvärk, och återhämtningen tar ofta lång tid. Det är viktigt att betona att det inte behöver vara fel på intellektet bara för att man är uttröttbar [1].
Sjuklig uttröttbarhet förekommer ofta efter traumatiska skall- och hjärnskador, hjärnhinneinflammation och stroke [2]. Det är när patienten blivit återställd från sina mer påtagliga handikapp och ska gå tillbaka till vardagsmiljön som besvären tydligt gör sig gällande och försvårar återgång till arbetet.
Denna sjukliga mentala uttröttbarhet är även mycket vanligt förekommande vid inflammatoriska sjukdomar i nervsystemet såsom multipel skleros [3], men den förekommer ibland även tidigt i utvecklingen av degenerativ sjukdom såsom en demenssjukdom eller Parkinsons sjukdom [4-6].
Uttröttbarheten behöver dock inte vara en följd av nervcellsskada. Även personer som lider av svår och långvarig värk besväras ofta av sjuklig mental uttröttbarhet, liksom personer som utsatts för långvarig och ogynnsam stress på arbetet eller genom personliga händelser [7, 8].
Det handlar kanske snarare om att hjärnans »underhållssystem« är drabbade.
Figur 1 illustrerar problemet sjuklig, mental uttröttbarhet och möjliga underliggande mekanismer, vilka belyses i denna artikel.
Identifiering och diagnostik
Även om klinisk anamnes och neuropsykologiska instrument kan användas för att bedöma vigilans/uthållighet och uppmärksamhet, saknar vi idag objektiva mätinstrument för att påvisa och gradera den mentala uttröttbarheten. Vi får inte heller någon vägledning vad gäller symtomens svårighetsgrad genom att titta på eller undersöka personen. Den som lider av mental uttröttbarhet ser frisk ut, beter sig även så och presterar ofta normalt i vanliga neuropsykologiska test. Besvärsgraden är dessutom beroende av omgivningens eller personens egna krav på intellektuell arbetsförmåga. I en lugn och för personen positiv miljö är den kognitiva kapaciteten kanske tillfredsställande, medan det i en miljö med mycket stimuli kan bli total »låsning« av hjärnfunktionerna.
Om man vill försöka förstå tillståndets natur skulle man kunna jämföra med allergi. Den allergiske har alltid sin allergi men besväras först då hon/han utsätts för allergen. På liknande sätt är det med uttröttbarhet; tillståndet finns där hela tiden, men besvären uppträder först då personen utsätts för stimulering och framför allt då ny information måste tas in och bearbetas.
Diagnostiken vilar i de flesta fall på rent kliniska grunder, och svårighetsgraden av besvären bedöms individuellt utifrån anamnesen.
Mental uttröttbarhet är inte någon specifik diagnos med diagnosnummer utan inryms tillsammans med emotionella och sensoriska symtom (Fakta) i diagnoserna posttraumatiskt eller postinfektiöst syndrom om besvären uppkommit efter skallskada eller infektion i nervsystemet. Om man använder den internationella sjukdomsklassifikationen ICD-10 återfinns den mentala uttröttbarheten med koncentrationssvårigheter i diagnosen »mild cognitive disorder« eller »neurasthenia«, och enligt den amerikanska psykiatriklassifikationen DSM-IV [9] inryms symtomen i »mild neurocognitive disorders«. Enligt Lindqvists och Malmgrens klassificering av organiska psykosyndrom [10] inryms uttröttbarheten inom det »asteno-emotionella syndromet«.
Svårt förklara hur sjuklig uttröttbarhet uppstår
När vi talar om komplexa system som hjärnan är det viktigt att utgå från identifierade referensramar. Referensramarna i denna artikel är kopplade till funktioner i hjärnan som associerar till underhålls- och infrastrukturella processer på cellulär nivå. De celler som engageras i dessa processer är också direkt involverade i informationsbehandlande processer; liknande funktionsstörningar och symtom kan förekomma i andra sammanhang än de vi refererar till här. Dessa berör vi endast i begränsad omfattning.
Trots de stora framsteg i kunskapen om hjärnans funktioner som gjorts, inte minst under senare år, är det mycket vi känner till endast i grova drag. Inte minst gäller detta kunskapen om hur våra mentala processer utförs. Här finns flera olika nivåer där man studerat hur hjärnan modulerar intryck, alltifrån molekylär nivå till funktionella studier av hjärnan.
Om vi utgår från en mer övergripande nivå kan vi dock identifiera funktioner som måste ingå. Den funktionella moduleringen av sensoriska inflöden har studerats avseende syn, hörsel och perception [11-13]. Det måste finnas funktioner som vi kan kalla mönsterigenkänning, beslutsfattande mm. Associerat till syninformationshantering finns tex särskilda enheter med neuronala kopplingar för igenkänning av räta linjer och horisontella och vertikala strukturer.
För andra signaltyper finns liknande enheter, men också andra mera anpassningsbara. Sådana enheter kan aktiveras omedvetet för att tex undertrycka ett bakgrundsljud och hindra det att passera upp till medvetandet. När fläkten – som varit igång hela dagen – stannar blir vi medvetna om att ljudet upphört och känner en tydlig lättnad. Detta är en indikation på att detta undertryckande varit förenat med en energikrävande process.
Enheter finns för olika funktioner och kan vara mer eller mindre dynamiska och formbara. I medvetna mentala processer rekryteras, formas och/eller aktiveras sådana enheter kontinuerligt. På samma sätt som för den omedvetna undertryckningen av fläktljudet är mentala processer, där sådana funktioner används, energikrävande. Vi har emellertid lättare att associera eventuell resulterande trötthet med dessa, eftersom de aktiveras i medvetna processer. När mentala funktioner skapats och formerats under normala betingelser kommer de att fungera optimalt under liknande betingelser. Om betingelserna ändras kan precisionen i funktionerna försämras, vilket leder till att osäkerheten i processerna ökar samtidigt med energiåtgången. Vi tittar närmare på detta längre fram.
Glutamatsignalering för intag och bearbetning av information
Hjärnan arbetar som en helhet, och störning av hjärnans normala arbetssätt minskar dess möjligheter att arbeta effektivt. Glutamatsignaleringen står för största delen av hjärnans excitatoriska aktivering. Resultatet av störning blir att hjärnan inte kan bearbeta lika stor informationsmängd på ett lika effektivt sätt som under normala förhållanden [14-16].
En logisk konsekvens skulle vara att hjärnan begränsar informationsintaget. Detta kan inte ske hur som helst. Information som varnar för fara måste kunna tas in och bearbetas för att trygga vår överlevnad.
Hur har naturen löst problemet att begränsa hjärnans informationsbearbetning utan att minska informationsintaget och utan att begränsa de intellektuella resurserna? Eftersom hjärnans glutamatsignalering är av stor betydelse för vår förmåga att ta in och bearbeta information, ska vi titta närmare på regleringen av glutamattransmissionen för att se om störning i denna signalering kan hjälpa till att förklara uttröttbarhet.
Nervcell aktiv i mental process – kan bli som muskelkramp
Låt oss först göra en ytterst förenklad beskrivning av vad som kan ske kring en nervcell som är aktiv i en mental process. Vi tänker oss att just denna nervcell vidarebefordrar signalen från en cell till en annan. För att göra detta är ett antal synapser på cellkroppen och/eller dendriterna aktiverade så att nervcellen är lämpligt polariserad för att motta en aktionspotential och föra den vidare.
I denna process kan ett stort antal synapser vara aktiva. Vid varje synaps finns astrocyter med uppgift att reglera glutamat- och jonnivåerna så att känsligheten för att skapa en ny aktionspotential blir den rätta [17-19]. Vid glutamatupptag kommer astrocytens volym primärt att öka genom osmos. Detta är en naturlig process, och normalt återgår volymökningen när glutamatet har stuvats undan. Om vi arbetar intensivt mentalt kommer emellertid flera närliggande nervceller att upprepat aktiveras, och återställningen hinner inte fullföljas innan nästa aktivering blir aktuell.
Astrocyterna kan rekrytera viss hjälp av närliggande celler, men om aktivitetsgraden är hög under längre tid kommer det inte att finnas mer hjälp att få eftersom transportmöjligheterna försämras på grund av minskat extracellulärt utrymme. Som ett resultat kommer polarisationsnivån att kontinuerligt vara hög, aktionspotentialer kommer att triggas spontant, och den mentala precisionen sjunker; vi känner oss trötta. En kort paus räcker vanligtvis för att återställa normalsituationen, och vi kan återuppta arbetet.
Nu kan emellertid situationen vara sådan att andra faktorer också påverkar reglerfunktionen. Astrocyterna har en roll också i de metabola processerna och i mera långsiktiga mentala specialiseringsprocesser. De metabola processerna prioriteras i allmänhet, och om (av någon anledning) delar av en tidigare adapterad specialiseringsprocess störts kommer denna att återskapas. Nu blir bilden annorlunda. I scenariot ovan ingår cellen i en adapterad verksamhet, tex en associationsbana.
Om denna process är störd kommer ny adaptation att ske parallellt med den avsedda aktiviteten, vilket resulterar i att mångfalt fler synapser aktiveras och kraven på astrocyternas arbetsförmåga mångfaldigas. Om många andra processer samtidigt har liknande problem blir astrocyternas uppgifter oöverstigliga. Vi får en utbredd cellsvullnad, och den viktiga jontransporten via extracellulärrummet störs (Figur 2). Vi får något som bildligt kan jämföras med kramp i en muskel.
Vi studerar nu de cellulära och biokemiska processer som är involverade.
Astrocyterna har central roll i glutamatsignaleringen
Glutamat signalerar från en nervcell till nästa genom att det frisätts från nervcellens presynaptiska terminal när en elektrisk impuls passerat axonet. Glutamat utövar därefter sina effekter på nervceller och omkringliggande gliaceller. För att specificiteten och effektiviteten i glutamatsignaleringen ska bli hög krävs att glutamat tas bort från synapsområdet omedelbart efter att det fullgjort sin signaleringsfunktion [20].
Det är astrocyterna som ansvarar för borttransporten av glutamat från det extracellulära utrymmet [19]. Astrocyterna tillhör hjärnans stödjeceller. De består av en cellkropp på ungefär 15 Ìm i diameter, från vilken långa utskott utgår. Efter färgning ser cellen ut som en stjärna i mikroskopet, därav namnet astrocyt eller astrogliacell. Utskotten sträcker sig mot synapsområden, nervcellskroppar och blodkärlens väggar och skapar kontakt med andra astrocytutskott så att cellerna ligger ordnade i stora cellförband [21-23].
Astrocyterna har välutvecklade transportproteiner för glutamat, vilka med hög aktivitet och specificitet tar upp glutamat från synapsområdet [24]. Dessa proteiner har isolerats från människa, där man benämnt dem excitatoriska aminosyratransportörer (excitatory amino acid transporters, EAAT 1–5). Motsvarande proteiner isolerade från råtta kallas glutamat– aspartattransportör (GLAST) [25] och glutamattransportör-1 (GLT-1) [26]. Nämnas kan att även nervcellerna har upptagssystem för glutamat [27], men dessa är inte lika effektiva som astrocyternas.
Väl inne i astrocyten omvandlas glutamat till glutamin, som inte har någon känd transmittorfunktion och som därför kan skickas tillbaka till nervcellen för att åter omvandlas till glutamat. Man har visat att astrocyterna frisätter glutamin när de registrerar hög nervcellstrafik i glutamatnervcellerna [28]. Genom detta arrangemang har naturen försäkrat en effektiv glutamatsignalering med hög precision och tålighet över tiden. Förutom upptagssystem för glutamat har astrocyterna också olika glutamatreceptorer, både jonotropa och metabotropa [22], så att även astrocyterna känner av glutamatmängden utanför cellerna.
GLT-1 viktigt transportprotein för extracellulärt glutamat
GLT-1 är det transportprotein som idag anses svara för det största glutamatupptaget. Det finns även hållpunkter för att GLT-1 verkar som en buffert som frisläpper en viss mängd glutamat, ibland ända upp till 35 procent, efter att det tagits upp från extracellulärrummet [29]. GLT-1 har således stora förutsättningar att reglera de extracellulära glutamatnivåerna och kan därmed påverka nervcellernas excitabilitetsnivå.
Figur 3 visar några mekanismer som reglerar bildningen av GLT-1 på astrocyterna.
Hjärnskada/infektion: astrocyterna klarar mindre glutamat
Man har i experimentella system visat att skada i nervsystemet leder till minskad förmåga hos astrocyterna att ta hand om glutamat [36-40]. Man har också påvisat nedreglering av GLT-1-uttrycket i hippocampus och hjärnbarken efter traumatiska hjärnskador [41, 42] och vid inflammatoriska och infektiösa tillstånd i nervsystemet [43-46].
Sådan minskad förmåga att ta hand om glutamat leder till att glutamathalterna runt nervcellerna ökar [47, 48]. Glutamat i hög koncentration under längre tid kan leda till nervcellsdöd genom att Ca2+ flödar in i nervcellerna via jonotropa glutamatreceptorer och startar program för celldöd [49]. Här koncentrerar vi oss i stället på hur neurotransmissionen kan påverkas långt innan nervcellsdöden inträder.
För att glutamattransmissionen ska ske med hög precision krävs att den extracellulära glutamatkoncentrationen inte överstiger några få Ìmol/l [20]. Vid störning i regleringen av extracellulärt glutamat ökar »bruset« i signaleringen, och precisionen i informationshanteringen minskar. Det är då rimligt att tänka sig att relativt sett fler signaler når upp till barken för bearbetning och identifiering, helt enkelt för att de avkännande systemen på lägre nivåer inte känner igen informationen och således inte kan avgöra dess natur. Den ständigt varierande informationen uppfattas som delvis ny, trots att så inte är fallet.
Vi kan ta ljudet från en lågfrekvent fläkt som exempel. Under normala förhållanden uppfattar man inte fläktljudet efter en stund, trots att det finns där. Hjärnan låter inte ljudimpulsmönstret nå upp till medvetandet – det »filtreras bort«. Detta är ändamålsenligt. Hjärnan skulle bli överbelastad om all information tilläts gå vidare upp till hjärnbarken. Vid ökat brus kan man se det som att man inte klarar att stänga av fläktljudet.
Hypotetiskt kan en störd reglering av de extracellulära glutamatnivåerna vara en neurobiologisk förklaring till den bristande »filtreringen« av information upp till hjärnans bearbetningscentra efter en hjärnskada.
Om mer information når hjärnbarken för bearbetning, kommer fler och större nervcellskretsar att aktiveras. Vid störd reglering av extracellulärt glutamat ökar de extracellulära glutamatnivåerna lokalt. Här måste dock betonas att den faktiska glutamatökningen sannolikt är högst begränsad när man ser till fall utan bakomliggande nervcellsskada. Under alla förhållanden kan det vara så att mekanismerna bakom mental uttröttbarhet kan vara bristande precision i transmissionen snarare än förhöjda extracellulära glutamatnivåer som en risk för eventuell celldöd.
Den störda regleringen av extracellulärt glutamat kan dock ge upphov till en rad effekter för transmissionen: astrocyterna ökar sin volym något [50], och som följd kommer extracellulära volymen att minska [51], vilket ger effekter på koncentrationen och transporten av neuroaktiva ämnen utanför nervcellerna och således på den extracellulära signaleringen mellan cellerna. Cellsvullnad leder också till att cellerna depolariseras något, och glutamatupptaget, som är spänningsberoende, minskar ytterligare. Relativ depolarisation leder dessutom till minskad kapacitet hos astrocyterna att ta hand om extracellulärt K+ som frisätts i transmissionen. Resultatet kan bli att de extracellulära K+-nivåerna ökar något, framför allt om transmissionen är intensiv.
Även måttligt förhöjda extracellulära K+-nivåer (8–10 mmol/l) har i experimentella system visats hämma nervcellernas frisättning av glutamat [52].
Vid störning av den extracellulära homeostasen är det troligt att mikrogliacellerna, en annan gliacellstyp, reagerar med bla bildning och frisättning av proinflammatoriska cytokiner som TNF(alfa) (tumörnekrosfaktor alfa) och interleukin6 (IL-6) [53]. Dessa cytokiner har visats ytterligare hämma astrocyternas kapacitet att ta hand om extracellulärt glutamat [44, 45].
Vid minskad glutamatupptagskapacitet hos astrocyterna minskar dessutom mängden tillgängligt glutamat inne i cellerna, varför glutaminbildningen minskar och sekundärt härtill även nybildningen av glutamat. Man vet också att glukosupptaget minskar, med störd metabolism som följd.
Vi ser här flera biokemiska underlag till minskad transmission, vilket skulle kunna vara det cellulära underlaget till uttröttbarheten.
Genom denna förklaringsmodell förstår vi varför personen klarar kognitivt krävande arbete en kort stund, medan längre tids stimulering skapar metabola förändringar på cellulär nivå som det tar lång tid att återställa. Vi ska även nämna att det finns andra teorier för uppkomst av uttröttbarhetstillstånd där genetiska, biokemiska, immunologiska, neuroendokrina och autonoma mekanismer och störningar, inklusive sömnstörningar, anses betydelsefulla [8, 54].
Stöd för vår förklaringsmodell
Vi vet idag att traumatisk skada, ischemi och begynnande degeneration är kopplade till inflammation med mikrogliaaktivering och cytokinproduktion i hjärnan. Intressant är att det även vid tillstånd utan uppenbar nervcellsskada, som depression, sömnbrist och sjukdomsbeteende, där den mentala uttröttbarheten ibland är mycket uttalad, förekommer en inflammatorisk reaktion i hjärnan med cytokinproduktion. Det är även känt från experimentella system att IL-1-tillförsel leder till minskad kapacitet för inlärning och minne. Även TNF(alfa) är direkt glutamatupptagshämmande för astrocyter.
Även vid smärttillstånd, där blod–hjärnbarriären är öppnad, har man påvisat inflammatorisk aktivitet med cytokinbildning i nervsystemet, och kliniskt vet vi att dessa patienter ofta lider av mental uttröttbarhet. Inflammation i hjärnan med mikrogliaaktivering och cytokinproduktion skulle kunna vara en orsak till mental uttröttbarhet, där en mekanistisk förklaring på det cellulära planet skulle kunna vara bristande glutamatreglering hos hjärnans astrogliaceller, eftersom vi vet att denna reglering är känslig för inflammatoriska mediatorer [46].
Sammanfattningsvis för vi fram en hypotetisk neurobiologisk förklaring till mental uttröttbarhet där störning i blod– hjärnbarriärfunktionen, inflammatorisk aktivitet med mikrogliaberedskap och cytokinbildning skulle kunna vara ett underlag för bristande astrogliareglering av glutamatsignaleringen.
Framtida bildbehandling på cellulär nivå kan ge mer kunskap
I enlighet med vad vi sagt tidigare och med vår nuvarande begränsade kunskap om hjärnans funktioner är det idag inte möjligt att slutligt visa om hypotesen är sann.
Bildbehandling på cellulär nivå kommer med all sannolikhet att i en framtid ge mer kunskap om cellulära mekanismer bakom mental uttröttbarhet.
Farmakologisk terapi kan bli möjlig
Patologisk uttröttbarhet kan ibland vara orsakad av oreparerbara skador på nervceller. Detta gäller tex för multipel skleros, stroke och traumatiska hjärnskador. Vid sådana tillstånd är det angeläget att behandling startas tidigt innan nya neurala kopplingar etableras [14, 18]. För att minska risken för sekundär oro och ångest, speciellt efter lättare traumatisk skada eller infektion, där uttröttbarhet förekommer utan nervcellsskador, är tidig information om tillståndets ofta godartade karaktär grundläggande.
Dessutom är det viktigt för personen att lära känna symtomen och sin egen begränsning med mål att undvika överbelastning av hjärnan och hamna i »total uttröttning«. Inflammationshämmande och cellavsvällande läkemedel skulle, om våra tankar visar sig riktiga, vara möjliga behandlingsvägar.
Om vår hypotes om uppkomsten av symtomen är korrekt handlar det om att minska bildningen av proinflammatoriska cytokiner och därigenom stärka den astrogliala glutamatupptagskapaciteten. Här kan xantiner och liknande ämnen [55] vara intressanta. Minocyklin, ett syntetiskt tetracyklinderivat, kan också vara av intresse, eftersom denna substans visats hämma mikrogliaaktivering och bildning av proinflammatoriska cytokiner [56].
Under senare år har substanser som ökar glutamatupptaget identifierats. Nicergolin [57], olika tillväxtfaktorer, en del lågmolekylära substanser såsom den hypofysära adenylatcyklasaktiverande peptiden PACAP (pituitary adenylate cyclase-activating polypeptide) [33, 58] och även metabotropa glutamatagonister [59] har visats kunna stimulera glutamattransporten i experimentella system, och de skulle kunna vara av intresse att vidareutveckla till produkter användbara på människa mot mental uttröttbarhet. Intressant nog har även AMPA-receptormodulatorer (AMPA=a-amino-3-hydroxi-5-metylisoxazol-4-propionsyra) visats kunna stärka kognitiva funktioner [60].
Mer direkta försök att stimulera GLT-1-uttrycket görs redan, och en intressant väg som nyligen rapporterats är via â-laktamer [61]. Upptäckten av splitsvarianter av GLT-1-proteinet kan vara en ledtråd som förklaring till varför uttröttbarheten blir kvar hos vissa människor även efter det att skadan eller sjukdomen är läkt. På längre sikt, när vi förstår mer om GLT-1-proteinets molekylära uppbyggnad, posttranslationella hopsättning och reglering i olika hjärnområden [62], skulle möjliga vägar för farmakologisk terapi kunna öppnas, även om andra terapiformer utifrån alternativa förklaringsmodeller till tillståndet kan visa sig utvecklingsbara, tex stresshantering, olika former av kognitiv terapi och andra farmakologiska behandlingsstrategier.
Med dagens ökande krav på mental kapacitet på den alltmer högteknologiska arbetsmarknaden kan mental uttröttbarhet bli ett svårt handikapp, inte minst eftersom detta handikapp är abstrakt för oss och inte »syns utanpå«. Det är därför av stor betydelse att vi intresserar oss för detta problem för att förstå uppkomstmekanismer som grund för effektiv behandling.
*
Potentiella bindningar eller jävsförhållanden: Inga uppgivna.
Summary
Cognitive impairment with decreased concentration capacity and mental fatigue makes rehabilitation from a stroke, brain trauma, meningitis or neuro-inflammation difficult, and often hampers going back to work. According to our hypothesis, down regulation of the glutamate transport protein GLT-1 in the hippocampus and cerebral cortex is one pathogenic factor underlying mental fatigue. Glutamate is one major signal substance for information intake and processing in the brain. The GLT-1 protein, localised to astrocytes and, upon activation, also microglial cells, is considered to play key roles in the regulation of extracellular glutamate. A slight disturbance of the astroglial regulation of extracellular glutamate could lead to local microglial activation with the production of proinflammatory cytokines, astrocyte swelling, and, due to such swelling, decreased extracellular space. During these conditions, there is a decrease in both the glutamate release and the neuronal transmission. It is proposed that this decreased transmission could be one correlate at the cellular level to the mental exhaustion that the person could experience. Pharmacological treatment, probably with glial cells as targets, could restore dysfunction in the glutamate transmission and make the person less susceptible to pathological mental fatigue.
Rönnbäck L, Persson M, Olsson T
Correspondence: Lars Rönnbäck, Institutionen för neurovetenskap och fysiologi, Sektionen för klinisk neurovetenskap och rehabilitering, Sahlgrenska Akademin vid Göteborgs Universitet, Guldhedsgatan 19, 1tr, SE-413 45 Göteborg, Sweden lars.ronnback@neuro.gu.se
FAKTA.
Vanliga symtom associerade med mental uttröttbarhet
Nedsatt koncentrationsförmåga
Subjektiva minnessvårigheter
Nedsatt simultankapacitet
Affektlabilitet
Humörinstabilitet
Ljud-/ljusöverkänslighet
Huvudvärk
REFERENSER
1.Azouvi P, Couillet J, Leclercq M, Martin Y, Asloun S, Rousseaux M. Divided attention and mental effort after severe traumatic brain injury. Neuropsychologia 2004;42:1260-8.
2.Kreutzer JS. Preface. Long-term outcome following severe brain injury. J Head Trauma Rehabil 2001;16:v-vi.
3.Krupp LB, Christodoulou C. Fatigue in multiple sclerosis. Curr Neurol Neurosci Rep 2001;1:294-8.
4.Herlofson K, Larsen JP. The influence of fatigue on health-related quality of life in patients with Parkinson´s disease. Acta Neurol Scand 2003;107:1-6.
5.Alves G, Wentzel-Larsen T, Larsen JP. Is fatigue an independent and persistent symptom in patients with Parkinson disease? Neurology 2004;63:1908-11.
6.Chaudhuri A, Behan PO. Fatigue and basal ganglia. J Neurol Sci 2000;179:34-42.
7.Hansson E, Rönnbäck L. Altered neuronal-glial signaling in glutamatergic transmission as a unifying mechanism in chronic pain and mental fatigue. Neurochem Res. 2004;29:989-96.
8.DeLuca J. (ed.) Fatigue as a Window to the Brain. 2005. A Bradford Book, The MIT Press, Cambridge, Massachusetts, London, England, 336pp.
9.American Psychiatric Association, Diagnostic and statistical menual of mental disorders, 4th ed. Washington DC: American Psychiatric Association, 1994.
10.Lindqvist G, Malmgren H. Organic mental disorders as hypothetical pathogenetic processes. Acta Psychiatr Scand 1993;88suppl373:5-17.
11.Buckner RL, Raichle ME, Miezin FM, Petersen SE. Functional anatomic studies of memory retrieval for auditory words and visual pictures. J Neurosci 1996;16:6219-35.
12.Petrovic P, Ingvar M. Imaging cognitive modulation of pain processing. Pain 2002;95:1-5.
13.Fox MD, Snyder AZ, Vincent JL, Corbetta M, van Essen DC, Raichle ME. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc Natl Acad Sci 2005;102:9673-78.
14.Dancause N, Barbay S, Frost SB, Plautz EJ, Chen D, Zoubina EV, Stowe AM, Nudo RJ. Extensive cortical rewiring after brain injury. J Neurosci 2005;25:10167-79.
15.Bütefisch CM. Neurobiological bases of rehabilitation. Neurol Sci 2006;27:S18-S23.
16.Carmichael ST. Cellular and molecular mechanisms of neural repair after stroke: Making waves. Ann Neurol 2006;59:735-42.
17.Hansson E, Rönnbäck L. Astrocytes in glutamate neurotransmission. FASEB J 1995;9:343-50.
18.Hansson E, Rönnbäck L. Glial neuronal signaling in the central nervous system. FASEB J 2003;17:341-348.
19.Hertz L, Zielke HR. Astrocytic control of glutamatergic activity: astrocytes as stars of the show. TRENDS in Neurosci 2004;27:735-43.
20.Yudkoff M, Nissim I, Daikhin Y, Lin ZP, Nelson D, Pleasure D, Erecinska M. Brain glutamate metabolism: neuronal-astroglial relationships. Dev Neurosci 1993;15:343-50.
21.Blomstrand F, Khatibi S, Muyderman H, Hansson E, Olsson T, Rönnbäck L. 5-Hydroxytryptamine and glutamate modulate velocity and extent of intercellular calcium signalling in hippocampal astroglial cells in primary cultures. Neuroscience 1999;88:1241-53.
22.Hansson E, Rönnbäck L. Astrocytic receptors and second messenger systems. Hertz L (ed) Advances in Molecular and Cell Biology 2004;31:475-501.
23.Abbott NJ, Rönnbäck L, Hansson E. Astrocyte-endothelial interactions at the blood-brain barrier. Nature Rev Neurosci 2006;7:41-53.
24.Diamond JS, Jahr CE. Synaptically released glutamate does not overwhelm transporters on hippocampal astrocytes during high-frequency stimulation. J Neurophysiol 2000;83:2835-43.
25.Storck T, Schulte S, Hofmann K, Stoffel W. Structure, expression, and functional analysis of a Na(+)-dependent glutamate/aspartate transporter from rat brain. Proc Natl Acad Sci USA 1992;89:10955-9.
26.Pines G, Danbolt NC, Bjoras M, Zhang Y, Bendahan A, Eide L, Koepsell H, Storm-Mathisen J, Seeberg E, Kanner BI. Cloning and expression of a rat brain L-glutamate transporter. Nature 1992;360:464-7.
27.Danbolt NC. Glutamate uptake. Prog Neurobiol 2001;65:1-105.
28.Bröer A, Deitmer JW, Bröer S. Astroglial glutamine transport by system N is upregulated by glutamate. Glia 2004;48:298-310.
29.Bergles DE, Tzingounis AV, Jahr CE. Comparison of coupled and uncoupled currents during glutamate uptake by GLT-1 transporters. J Neurosci 2002;1:10153-62.
30.Björklund U, Persson M, Ängehagen M, Rönnbäck L, Hansson E. Characterization ofneuron enriched cultures, grown in defined media, show a possible connection between GLT-1 expression and presence of VGLUT1 positive neurons. Manuscript.
31.Perego C, Vanoni C, Bossi M, Massari S, Basudev H, Longhi R, Pietrini G. The GLT-1 and GLAST glutamate transporters are expressed on morphologically distinct astrocytes and regulated by neuronal activity in primary hippocampal cocultures. J Neurochem 2000;75:1076-84.
32.Levy LM, Lehre KP, Walaas SI, Storm-Mathisen J, Danbolt NC. Down-regulation of glial glutamate transporters after glutamatergic denervation in the rat brain. Eur J Neurosci 1995;1:2036-41.
33.Figiel M, Engele J. Pituitary adenylate cyclase-activating polypeptide (PACAP), a neuron-derived peptide regulating glial glutamate transport and metabolism. J Neurosci 2000;20:3596-605.
34.Aronica E, Gorter JA, Ijlst-Keizers H, Rozemuller AJ, Yankaya B, Leenstra S, Troost D. Expression and functional role of mGluR3 and mGluR5 in human astrocytes and glioma cells:opposite regulation of glutamate transporter proteins. Eur J Neurosci 2003;17:2106-18.
35.Figiel M, Maucher T, Rozyczka J, Bayatti N, Engele J. Regulation of glial glutamate transporter expression by growth factors. Exp Neurol 2003;183:124-135.
36.Rothstein JD, Van Kammen M, Levey AI, Martin LJ, Kuncl RW. Selective loss of glial glutamate transporter GLT-1 in amyotrophic lateral sclerosis. Ann Neurol 1995;38:73-84.
37.Knecht K, Michalak A, Rose C, Rothstein JD, Butterworth RF. Decreased glutamate transporter (GLT-1) expression in frontal cortex of rats with acute liver failure. Neurosci Lett 1997;229:201-3.
38.Li S, Mallory M, Alford M, Tanaka S, Masliah E. Glutamate transporter alterations in Alzheimer disease are possibly associated with abnormal APP expression. J Neuropathol Exp Neurol 1997;56:901-11.
39.Hazell AS, Rao KV, Danbolt NC, Pow DV, Butterworth RF. Selective down-regulation of the astrocyte glutamate transporters GLT-1 and GLAST within the medial thalamus in experimental Wernicke´s encephalopathy. J Neurochem 2001;78:560-8.
40.O´Shea RD. Roles and regulation of glutamate transporters in the central nervous system. Clin Exp Pharmacol Physiol 2002;29:1018-23.
41.Rao VL, Baskaya MK, Dogan A, Rothstein JD, Dempsey RJ. Traumatic brain injury down-regulates glial glutamate transporter (GLT-1 and GLAST) proteins in rat brain. J Neurochem 1998;70:2020-7.
42.Yi JH, Pow DV, Hazell AS. Early loss of the glutamate transporter splice-variant GLT-1v in rat cerebral cortex following lateral fluid-percussion injury. Glia 2005;49:121-133.
43.Szymocha R, Akaoka H, Dutuit M, Malcus C, Didier-Bazes M, Belin MF, Giraudon P. Human T-cell lymphotropic virus type 1-infected T lymphocytes impair catabolism and uptake of glutamate by astrocytes via Tax-1 and tumor necrosis factor alpha. J Virol 2000;74:6433-41.
44.Fine SM, Angel RA, Perry SW, Epstein LG, Rothstein JD, Dewhurst S, Gelbard HA. Tumor necrosis factor alpha inhibits glutamate uptake by primary human astrocytes. Implications for pathogenesis of HIV-1 dementia. J Biol Chem 1996;271:15303-6.
45.Su ZZ, Leszczyniecka M, Kang DC, Sarkar D, Chao W, Volsky DJ, Fisher PB. Insights into glutamate transport regulation in human astrocytes: cloning of the promoter for excitatory amino acid transporter 2 (EAAT2). Proc Natl Acad Sci USA 2003;100:1955-60.
46.Rönnbäck L, Hansson E. On the potential role of glutamate transport in mental fatigue. J Neuroinflammation 2004;1:22.
47.Torp R, Lekieffre D, Levy LM, Haug FM, Danbolt NC, Meldrum BS, Ottersen OP. Reduced postischemic expression of a glial glutamate transporter, GLT-1, in the rat hippocampus. Exp Brain Res 1995;103:51-8.
48.Legay V, Deleage C, Beaulieux F, Giraudon P, Aymard M, Lina B. Impaired glutamate uptake and EAAT2 downregulation in an enterovirus chronically infected human glial cell line. Eur J Neurosci 2003;17:1820-8.
49.Choi DW. Excitotoxic cell death. J Neurobiol 1992;23:1261-76.
50.Hansson E. Metabotropic glutamate receptor activation induces astroglial swelling. J Biol Chem. 1994;269:21955-61.
51.Sykóva E. Glial diffusion barriers during aging and pathological states. Prog Brain Res. 2001;132:339-63.
52.Meeks JP, Mennerick S. Selective effects of potassium elevations on glutamate signaling and action potential conduction in hippocampus. J Neurosci. 2004;24:197-206.
53.Persson M, Brantefjord M, Hansson E, Rönnbäck L. Lipopolysaccharide increases microglial GLT-1 expression and glutamate uptake capacity in vitro by a mechanism dependent on TNF-. Glia 2005;51:111-120.
54.Rönnbäck L. Fatigue (review of ÒFatigue as a window to the brainÓ ed by DeLuca J issued in Clinical and Cognitive Neuropsycology 2005;336pp Cambridge, Mass, MT Press) JAMA 2006;295:444-445.
55.Schubert P, Rudolphi K. Interfering with the pathologic activation of microglial cells and astrocytes in dementia. Ahlzheimer Dis Assoc Disord 1998;12 suppl. 2:S21-S28.
56.Tikka T, Fiebich BL, Goldsteins G, Keinanen R, Koistinaho J. Minocycline, a tertracycline derivative, is neuroprotective against excitotoxicity by inhibiting activation and proliferation of microglia. J Neurosci 2001;21:2580-8.
57.Nishida A, Iwata H, Kudo Y, Kobayashi T, Matsuoka Y, Kanai Y, Endou H. Nicergoline enhances glutamate uptake via glutamate transporters in rat cortical synaptosomes. Biol Pharm Bull 2004;27:817-20.
58.Figiel M, Maucher T, Rozyczka J, Bayatti N, Engele J. Regulation of glial glutamate transporter expression by growth factors. Exp Neurol 2003;183:124-35.
59.Aronica E, Gorter JA, Ijlst-Keizers H, Rozemuller AJ, Yankaya B, Leenstra S, Troost D. Expression and functional role of mGluR3 and mGluR5 in the human astrocytes and glioma cells: opposite regulation of glutamate transporter proteins. Eur J Neurosci 2003; 17:2106-18.
60.Lynch G. AMPA receptor modulators as cognitive enhancers. Curr Opin Pharmacol 2004;4:4-11.
61.Rothstein JD, Patel S, Regan MR, Haenggeli C, Huang YH, Bergles DE, Jin L, Dykes Hoberg M, Vidensky S, Chung DS, Toan SV, Bruijn LI, Su ZZ, Gupta P, Fisher PB. Beta-lactam antibiotics offer neuroprotection by increasing glutamate transporter expression. Nature 2005;433:73-7.
62.Kalandadze A, Wu Y, Robinson MB. Protein kinase C activation decreases cell surface expression of the GLT-1 subtype of glutamate transporter. Requirement of a carboxyl-terminal domain and partial dependence on serine 486. J Biol Chem 2002;277:45741-50.

Illustration: Eva Kraft, Göteborg
Figur 1. Illustration av uttröttbarhetstillstånd med koncentrations- och minnessvårigheter. Om personen utsätts för mycket stimuli blir hon/han helt slut och orkar inte fortsätta med arbetet. Vi tänker oss att större delar av hjärnan än normalt aktiveras (mitten). I bildens nedre del ger vi en förklaring till besvärens uppkomst på cellnivå. Störning av regleringen av glutamathalten (Glu) utanför nervcellerna kan leda till astrocytsvullnad och minskat extracellulärt utrymme. Under sådana förhållanden minskar glutamatsignaleringen, vilket kan vara ett korrelat till den mentala uttröttbarhet personen upplever.
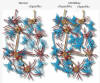
Figur 2. Till vänster: Förenklad illustration av neuronalt nätverk bestående av nervcellerna A-G (röda) under normala förhållanden. Cell A aktiverar (markeras med gul färg) B synaptiskt, varefter C och senare D aktiveras. Omgivande astrocyter (blå) känner av nervcellsaktiviteten och interagerar med nervcellerna. Resultatet blir att ett »specifikt« svar förs vidare. Till höger: Efter en hjärnskada ses en »sprouting«, vilken resulterar i att nervcell C aktiverar både D och E. Genom att glutamat »spills över«, och på grund av att den extracellulära glutamathalten är förhöjd, aktiveras cell G, vilken i sin tur aktiverar cell B. Resultatet blir aktivering av större neuronala kretsar, astroglial svullnad och »ospecifik« signalering i tillägg till den »specifika«. Bruset är således något ökat. Den ökade svullnaden av gliacellerna bidrar dessutom till att förstärka och bibehålla dessa processer genom begränsning av det extracellulära utrymmet.

Figur 3.I cellkultur har man sett att astrocyterna bildar GLT-1 i närvaro av glutamatnervceller [30]. Det verkar som om ämnen från nervcellerna stimulerar till GLT-1-bildningen, och nervcellstrafik aktiverar ytterligare GLT-1 [31]. GLT-1 har å andra sidan visats nedregleras efter glutamaterg denervation [32]. GLT-1 står också under kontroll av hypotalamus och hypofysen genom peptiden PACAP (pituitär adenylatcyklasaktiverande peptid) [33], som bildas i nervceller i hypofysen. GLT-1-uttrycket kan även regleras av astrogliacellerna själva. Genom att aktivera glutamatreceptorerna mGluR3 respektive mGluR5 har man visat att GLT-1-uttrycket ökar respektive minskar på cellerna [34]. Även ett antal tillväxtfaktorer såsom EGF (epidermal growth factor), TGF(alfa) (transforming growth factor-alpha), FGF-2 (fibroblast growth factor-2) och PDGF (platelet derived growth factor) ökar bildningen av GLT-1 [35].